 Vers
la page d'accueil
d'OPIE-Insectes
Vers
la page d'accueil
d'OPIE-Insectes
Vers
la page d'accueil
d'OPIE-Insectes
Three Successive Regulation Patterns of Gypsy Moth (Lepidoptera, Lymantriidae) Populations in the Mamora Cork Oak Forest (Morocco)
Alain FRAVAL and Claire VILLEMANT
fraval@paris.inra.fr,villeman@cimrs1.mnhn.fr
ABSTRACT
The gypsy moth, Lymantria dispar (L.) is a widespread forest defoliator throughout the northern hemisphere. In Morocco, pest populations went through cyclic outbreaks in many cork oak forests (Quercus suber L., Fagaceae). During a 20-yr. observation period (1972-1991), the population was regulated in three successive ways. The first one took place during a 10-yr. gradation period and the others occurred later during a localized outbreak.
1. Trophic interactions between L. dispar and its host-plant caused the pest population to collapse: after 2 or 3 years of increasing population density, the pest rapidly entirely defoliated the oaks so that almost all larvae starved to death (no virus epizootic occurred). In the summer, a few weeks after defoliation, the cork oaks produced new leaves but no new foliage appeared in the following spring. Surviving gypsy moth individuals were compelled to feed on tough foliage with poor food value and died.
2. Parasitoid wasps attacked young gypsy moth larvae and greatly decreased the number of pests so that oaks were not defoliated. Parasitoids of old larvae and pupae eliminated most of the surviving pest individuals.
3. Predators and destroyers of the gypsy moth egg masses ate the eggs or scattered them far from the food source so that only a few pest individuals survived.
Introduction
The gypsy moth, Lymantria dispar (L.) (Lep., Lymantriidae) is one the most significant forest pests. Its distribution area extends throughout the northern hemisphere, from Japan where larvae exploit larches (Larix spp.) to the western United States. In North Africa, as in North America and Europe, gypsy moths defoliate the hardwood forests, mostly oaks (Quercus spp.). In Morocco, the pest is present in every cork oak, Quercus suber (L.), and holm oak, Q. ilex (L.) forests. It also attacks deciduous oaks such as Q. pyrenaica (Willd.) and fruit trees (Prunus spp.). The gypsy moth was first mentioned in 1919 in the Mamora forest, near Rabat. A few years later, de Lépiney (1930) studied its biology and mortality factors.
New research has been undertaken since 1972: it was directed towards describing gypsy moth population dynamics, improving sampling methods and experimentally introducing new infectious agents (Hérard & Fraval, 1980 ; Fraval, 1984 ; 1986 ; 1989 ; Fraval & Villemant, 1995; Graf et al., 1989 ; Villemant, 1989).
Behavior and population dynamics of the gypsy moth are well known only in cork oak forests located along the Atlantic coast (from Tangier in the north to Casablanca in the south). In these landscaped forests, Q. suber stands are pure and all trees are of the same age. The trees are old and many of them are damaged. Undergrowth shrubs which vary greatly in density are unpalatable for the gypsy moth larvae (Villemant & Fraval, 1998).
Gypsy moth populations have several important features. Females are not able to fly. Eggs are laid on vertical or overhanging supports, usually cork trunks, and never on the forest floor. The entomophagous insect complex has a low diversity and is not very effective. There is only one oophagous parasitoid, Ooencyrtus kuvanae (Howard) (Hym., Encyrtidae) which was introduced in Morocco between 1927 and 1928 (de Lépiney, 1932). No epizootic occurs during the pest outbreaks and the nuclear polyhedrosis virus is notably absent. Predators and destroyers of the gypsy moth egg masses are conspicuous mortality factors. They destroy eggs by predation and by disrupting the surface of egg masses so that fragments of dislocated egg clusters are thrown down to the ground by the wind. In the Atlantic cork oak forest conditions, most of the eggs that have fallen are eaten by predators and only a few of them hatch. However, young larvae hatching on the forest floor are unable to reach the oak's foliage (Fraval, 1986; Villemant & Fraval, 1992).
In late June and early July, larvae of Dermestes lardarius (L.) (Col., Dermestidae) attack newly laid egg masses. Larvae of Tenebroides maroccanus (Reitter) (Col., Trogossitidae) then excavate egg masses, disrupting their cohesiveness and predating many eggs. During July, larvae of the Dermestidae Trogoderma versicolor meridionalis (Kraatz) and Anthrenus vladimiri Menier & Villemant (1993) also penetrate egg masses. In winter and early spring, lepidopteran larvae of Niditinea fuscipunctella (Haworth) (Tineidae) and Aglossa caprealis (Hübner) (Pyralidae) destroy many egg masses (Villemant, 1993; Menier & Villemant, 1993; Villemant & Andreï-Ruiz, 1999).
Gypsy moth outbreaks in the Mamora forest are spectacular. Larvae completely defoliate significant areas of forest which often cover tens of thousands of hectares. Such extensive defoliation has never been observed before Morocco became a protectorate and forests were managed (towards 1912). After violence had receded due to security measures, the forest was grazed and fires, which regularly broke out late in spring (when gypsy moth larvae were present), stopped. Since that time, defoliated areas were mapped each year. These maps show that a short latency period (with no defoliation at any site) alternates with several years of pest outbreak. The gypsy moth outbreaks seemed to shift first eastward and then westward. Identical cyclical gradation, with a periodicity of about 11 years, occurs synchronically in several other cork oak forests. The distance between Mamora forest and these much smaller forest stands never exceeds 100 km. In other cork oak forests (apparently similar and with the same climate), gypsy moth populations remain latent. Large populations are never or, at the most, very occasionally observed.
In the Mamora forest, the latest gypsy moth outbreak began in 1972 and ended in 1982. Variations in the coverage and location of defoliated areas were probably related to the dispersal behavior of the insect (fig. 1). The prolonged outbreak phase seems to result from the juxtaposition of asynchronized localized outbreaks.
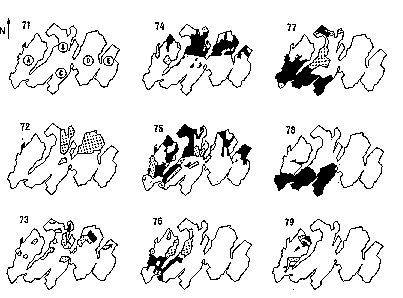
Figure 1: Map of gypsy moth defolation areas in the Mamora forest
from 1971 to 1979
A to E : cantons ; black surface : complete defoliation ; dotted surface
: partial defoliation
Owing to regular observations (from 1976 to 1982) along a transect, the mechanism leading to localized gradations could be described. The main factor affecting population level fluctuation was the trophic interaction between the pest and its host-plant. Since 1986, after an unusually prolonged latency period, a localized gradation broke out in the northwestern part of the forest (Canton A). Defoliation remained at a low level and did not extend to any other forest areas. The decrease in gypsy moth populations was induced by larvae and pupae parasitoids in 1986-1987 and 1987-1988, and by oophagous insects in 1988-1989 and 1989-1990 (Fraval et al., 1988; Villemant & Ramzi, 1995).
In this paper we give details of all these population regulation patterns, after describing the experimental survey that was designed for studying gypsy moth population dynamics. The first regulation pattern (trophic regulation) is the most common and was already described by Fraval (1984 ; 1986).
Experimental setup
A 7 km long permanent transect (fig. 2) runs from the forest house of El Menzeh to trench B1 (northwestern part of canton B). It was composed of 57 aligned plots of 10 trees each. Plots were 100 m apart. Observations were made on each tree from 1976 to 1982. Gypsy moth individuals were counted on trunks and in crowns from ground level. Every year, egg masses were counted and their destroyed surface was evaluated. Later in the season, larvae and egg laying females were numbered each week. Pupae were not counted because they cannot be easily distinguished from remaining dead pupae of the previous generation.
Figure 2: Principal experimental setups established in the western
part of the Mamora forest for studying gypsy moth population
dynamics
1 : transect (7 km) ; 2 : network (covers all of canton A) ; 3 : El
Hanchat (4 ha) ; black triangles : forest houses
The proportion of each egg mortality factor was evaluated in the laboratory by counting eggs (parasitized, predated, unfertilized or with a dead embryo). Larvae and pupae sampled in the field were cultured on cork oak foliage to assess the impact of parasitoids and any epizootics present (Fraval et al., 1980). Data were analyzed by several statistical methods. Raw data and more comprehensive variables obtained by multivariate procedures and hierarchic classification were plotted graphically. Many reasons besides practicality justified the use of a transect: the prospected area was sufficiently large to exhibit several patterns of gypsy moth population evolution, and comprehensive and meaningful graphics could be easily obtained (Fraval, 1983).
Another setup, the " network " was designed and established in 1978 in order to monitor the evolution of pest population density on a 20 000 ha forest area (canton A). This made it easier to forecast defoliation and take appropriate action. 240 plots of 40 trees each were more or less evenly distributed, about 1 km apart from each other (one plot to each 100 ha). Because of the scale of the network and the fact that available means were limited, observations and sampling could not be carried out weekly. Generally, cork oak foliage conditions and defoliation rates were estimated from an airplane. Data were processed and mapped by an in-house computer program since 1987 (Fraval, 1987 unpublished).
In 1987, at the site of El Hanchat near the town of Kenitra (canton A), a third setup was established in order to study a very localized (4 ha) outbreak. The spatial distribution of egg masses, the oophagous impact and the change in foliage were recorded on 143 adjacent cork oaks. Spatial patterns of the gypsy moth population were described (Fraval et al., 1988; Villemant & Ramzi, 1995).
Regulation pattern I
Figure 3 illustrates the various regulation patterns of gypsy moth population by showing the quantitative and qualitative changes affecting cork-oak foliage during a 4 yr. period (Villemant & Fraval, 1995). The first regulation pattern appears at the top of the diagram.
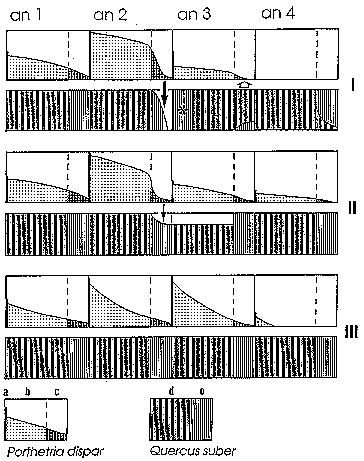
Figure 3: The three regulation patterns of gypsy moth populations in the
Mamora forest
I : regulation by lack of food and cork oak reaction ; II : regulation
by parasitoids ; III : regulation by oophagous predators.
Lymantria dispar : a (number of eggs laid in July) ; b (living eggs)
; c (larvae, pupae and adults). Quercus suber : d (old foliage, unfavourable)
; e (new foliage, favourable).
Black arrow : defoliation ; asterisk : summer renewing of the foliage
; white arrow : phenological disruption.
Initially (first year), there are few gypsy moths (about 1 000 to 100 000 eggs per tree) and larvae feed exclusively on new leaves because in normal conditions cork oaks in the Mamora forest renew all of their foliage in spring. The defoliation rate is low and well-fed individuals produce numerous offspring.
In spite of a relatively widespread mortality by egg predation and parasitism, larvae of the second generation hatch in large numbers during a relatively short period (about 1 month). Larvae rapidly consume all the new cork oak leaves. After a week of inactivity, they simultaneously attack the old cork oak foliage and other plants such as Chamaerops humilis L. present in the vicinity. All of this foliage is tough and indigestible and almost all the larvae die of starvation. Only a few individuals pupate and produce the third generation.
Eggs of this new generation are partly sterile. The parasitoid O. kuvanae and oophagous predators destroy most of the living eggs. In addition, many embryos die before hatching mainly as a result of the parents' malnutrition. Consequently, very few larvae actually hatch. In reaction to the defoliation during the previous year, cork oaks which produced new leaves in July do not renew their foliage in the following spring. At that time, larvae which have to eat old remaining foliage cannot develop. The gypsy moth population collapses and during the following years, cork oaks revert back to renewing all of their foliage in spring .
Regulation pattern II
The starting point is the same. Climatic conditions in the autumn and winter induce a prolonged hatching period during the following spring: a little over two months pass between the hatching of the first and the last larvae. Several hymenopteran parasitoid generations may follow each other. Notable among these are Braconidae - Cotesia melanoscela (Ratzeburg), Glyptapanteles porthetriae (Müsebeck) and Meteorus pulchricornis Wesmael - which live on young larvae, and Brachymeria intermedia Nees (Chalcididae), a pupal parasitoid. Mortality due to the parasitoids has a conspicuous effect on gypsy moth population levels and defoliation is not complete. Mortality of lately hatched larvae is also significant because they feed on older and scarcer foliage. Consequently few gypsy moths ever reach adulthood.
The number of gypsy moths at the start of the third generation is low. Most individuals have been destroyed by oophagous insects or larval parasitoids which remain relatively numerous. The following year, the same mortality factors occur and the gypsy moth population collapses.
Regulation pattern III
The start of the gradation is still the same. Eggs are conspicuously destroyed by oophagous predators because increasing population levels of these enemies were favored by specific conditions affecting the forest site. Damaged trees provide numerous shelters and food resources under bark flaps (Villemant, 1993; 1995; Villemant & Ramzi, 1995).
Only a few larvae hatch and the defoliation rate is low. Well-fed individuals produce an abundant progeny. Eggs of the third generation are numerous but most of them die owing to a significant predator activity. All remaining larvae are destroyed by parasitoids (which always occurs in very low level populations).
Discussion
Gypsy moth population regulation by cork oak feedback (pattern I) has regularly occurred in the Mamora forest during the pest gradation from 1972 to 1982. The same regulation pattern was shown in other cork oak forests along the Atlantic coast (Khemis Sahel and Aïn Felfel) and in the Rif (Bou-Hani). However it was not observed by de Lépiney (1930) when he studied gypsy moth population dynamics in the Mamora forest. Its occurrence may depend on current specific forest conditions (age and health of trees, climatic conditions) which lead to cork oaks renewing their foliage every year. In other neighboring cork oak forests, trees do not renew their leaves every year and gypsy moth populations remain stable (Fraval & Villemant, 1998). In other parts of the Mediterranean country, the pest population rarely collapses because of starvation and this mode of regulation is often considered to be of minor importance (Luciano & Prota, 1984).
Regulatory feedback by lack of food seems to be relatively unusual. It has been suspected in several host-plant insect interactions such as between L. dispar and Quercus velutina Lam. or between L. dispar and Betula populifera Marsh (Wallner & Walton, 1979). The best known cases of this regulation mechanism are:
- defoliation of Betula pubescens Erlich by Epirrita automnalis Bkh. (Lep. Geometridae) induces host-plant resistance to the pest. Two mechanism (of unknown origin) occur : the first of these takes place almost immediately and limits development of pest offspring, the second one induces a prolonged resistance (Haukioja, 1982);
- populations of the large bud moth, Zeirapheira diniana Genée (Lep. Tortricidae), increase in favorable conditions until larch foliage becomes indigestible. Starvation then induces decline in the pest population (Baltensweiler et al., 1977).
All authors noted that natural enemies play a major role in the decline of gypsy moth populations. In most cases, epizootics are the main mortality factor. In Morocco however, pathogens are unknown and pest mortality depends principally on two entomophagous guilds : egg mass predators and destroyers as well as larval or pupal parasitoids. The oophagous predators guild seems to be of major importance in Moroccan cork oak forests owing to conspicuous species diversity and their impact (Villemant & Ramzi, 1995). The activity of egg predators is quite regular. Together with the egg parasite O. kuvanae (which alone never has a conclusive impact), they sometimes destroy up to 90% of the eggs (50% on average). Larval and pupal parasitoids in Morocco are not specific to gypsy moths. They are generalist parasitoids and are found in all cork oak forests even when gypsy moth population levels remain constantly low (Villemant & Andreï-Ruiz, 1999).
Our observations have shown that entomophagous insects have a major impact on localized outbreaks occurring during a prolonged gypsy moth latency period (for the forest as a whole). In Mamora, a population decrease caused by entomophagous insects was already locally observed by de Lépiney (1930) and at the beginning of the next pest gradation (1972-1982) by Hérard and Fraval (1980).
In addition to the constantly low gypsy moth populations levels observed in some forests (Témara, Larache), gypsy moth populations in Moroccan cork oak forests can be regulated in one of three ways. Two of these are highly original : on the one hand, populations may decline because of shortage and qualitative modifications of the foliage conditioned by pest density pressure and climatic factors (Fraval, 1986), on the other hand the decline may be induced by the activity of oophagous predators.
Changes affecting gypsy moth populations were forecast effectively during the last gradation study by using a trophic regulation model (Fraval, 1986). Later, this model was unable to explain the behavior of localized outbreaks. Luckily, pest density measurements which are necessary for deciding on control action provide overvaluation so that the risk of overlooking a potential outbreak is quite small.
Since 1991 supervision over the main experimental setup has been stopped. Further wide scale investigations would probably be of little value anyway, because cork oak stands grow older and are not regenerated. Forest health worsens with illegal cork oak stripping and cutting and with increasing damage by xylophagous organisms: the fungus Diplodia mutila (Fri.) Mont. (Coelomycetes) (Bakry et al., 1999) and coleopteran insects including Cerambyx cerdo L. (Cerambycidae) and Platypus cylindrus (Platypodidae) (Villemant & Fraval, 1993; El Antry, 1999). Therefore, conditions affecting gypsy moth populations will probably differ considerably from those already observed.
REFERENCES CITED
Bakry M., El Antry S., Satrani B. & Oubrou, 1999. Réaction de différentes provenances de chêne-liège à l'action pathogène de Diplodia mutila. IOBC/WPRS Bull. (sous presse)
Baltensweiler W., Benz G., Bovey P. & Delucchi V., 1977. Dynamics of large bud moth population. Annu. Rev. Entomol., 22 : 79-100.
El Antry S., 1999. Biologie et dégâts de Cerambyx cerdo mirbecki Lucas (Coléoptère, Cerambycidae) en subéraie de la mamora (Maroc). IOBC/WPRS Bull. (sous presse)
Fraval A., 1983. Echantillonnage de Lymantria dispar en subéraie marocaine atlantique. In S. Frontier : Stratégies d'échantillonnage en écologie animale. Paris : Masson : 273-290.
Fraval A., 1984. Influence de la qualité et de la quantité de l'alimentation sur les fluctuations des populations de Lymantria dispar (L.) (Lep. Lymantriidae) en forêt de la Mamora (Maroc). Agronomie, 4 (9) : 819-828.
Fraval A., 1986. La régulation des populations de Lymantria dispar (L.) (Lép. Lymantriidae) en subéraie marocaine atlantique: effets du climat, des entomophages et des facteurs anthropiques. Z. angew. Entomol., 102 (1) : 38-52.
Fraval A., 1989. Dynamique des populations. In A. Fraval : Lymantria dispar. Rabat : Actes Editions (Documents scientifiques et techniques) : 53-72.
Fraval A. & Villemant C., 1995. La lutte biologique contre Le Bombyx disparate Porthetria dispar (L.) (Lép. Lymantriidae) au Maroc. IOBC/WPRS Bull., 18 (6) : 83-86.
Fraval A. & Villemant C., 1998. La Mamora et ses ennemis. Les dossiers de l'environnement de l'INRA. N° 15 Forêts : 133-146.
Fraval A., Questienne P. & Jarry M., 1980. Démographie de Lymantria dispar (L.) en subéraie de la Mamora. Etude le long d'un transect : 1976-1977-1978. Ann. Rech. for. Maroc, 20 : 3-64.
Fraval A., Villemant C., Chorfi B. & Ramzi H., 1988. Etude des structures spatiales d'une infestation locale de la subéraie de la Mamora par le Bombyx disparate, Lymantria dispar (L.) (Lép. Lymantriidae), en 1987. Ecol. mediterr., 14 (3-4) : 101-119.
Graf P., Hamdaoui M., Ramzi H. & Fraval A., 1989. Surveillance et lutte. In A. Fraval : Lymantria dispar. Rabat : Actes Editions (Documents scientifiques et techniques) : 77-92.
Haukioja E., 1982. Inducible defences of white birch to a geometrid defoliator, Epirrita automnata. Proc. 5th. Int. Symp. Insect Plant relatioships, Wageningen : Pudoc : 199-203.
Hérard F. & Fraval A., 1980. La répartition et les ennemis naturels de Lymantria dispar (L.) (Lep. Lymantriidae) au Maroc; 1973-1975. Acta Oecol., Oecol. appl., 1(1) : 35-48.
Lépiney J. de, 1930. Contribution à l'étude du complexe biologique de Lymantria dispar L. Mém. Soc. Sci. nat. Maroc, 23, 66 p.
Lépiney J. de, 1932. Le rôle de la Direction des Eaux et Forêts du Maroc et de l'Institut Scientifique Chérifien dans la lutte biologique entreprise contre Lymantria dispar L. à l'aide de Schedius kuwanae How. In : C. R. Ve Cong. int. Entomol., Paris, 5 : 807-812.
Luciano P. & Prota R., 1984. Osservazioni su alcuni fattori che influenzano la dinamica i popolazione di Lymantria dispar L. In : 4e Simp. Dinamica Populazioni, Parme, 22-24 oct. 1981 : 125-139.
Menier J.J. & Villemant C., 1993. Description et biologie de Anthrenus (Peacockia n. sgen.) vladimiri, n. sp. prédateur des pontes de Porthetria dispar (L.) au Maroc [Coleoptera, Dermestidae ; Lepidoptera Lymantriidae]. Rev. fr. Entomol. (N.S.), 15 (2) : 61-66.
Villemant C., 1989. Ennemis naturels et introduits des oeufs et des pontes. In A. Fraval : Lymantria dispar. Rabat : Actes Editions (Documents scientifiques et techniques) : 93-124.
Villemant C., 1993. Les prédateurs-démanteleurs des pontes de Porthetria dispar dans les subéraies marocaines. Thèse Univ. Orléans. Vol 1 : 267 p., vol 2 (annexes) : 91 p.
Villemant C., 1995. Les prédateurs-démanteleurs des pontes de Porthetria dispar (L.) (Lép. Lymantriidae) en subéraie marocaine. IOBC/WPRS Bull., 18 (6) : 87-94.
Villemant C., 1998. Les prédateurs-démanteleurs des pontes de Lymantria dispar en subéraie de la Mamora (Maroc). Bull. Soc. zool. Fr., 123 (1) : 75-81.
Villemant C. & Fraval A., 1992. Les ennemis des oeufs de Porthetria (Lymantria) dispar (L.) (Lép. Lymantriidae) au Maroc : inventaire et problèmes relatifs à l'évaluation de leur impact. Bull. Inst. sci. (Rabat), 16 : 160-172.
Villemant C. & Fraval A., 1993. La faune entomologique du chêne-liège en forêt de la Mamora (Maroc). Ecol. mediterr., 19 (3/4) : 89-98.
Villemant C. & Fraval A., 1995. Le Bombyx disparate Porthetria dispar (L.) (Lép. Lymantriidae) dans la subéraie de la Mamora (Maroc) : trois modes successifs de régulation des populations. IOBC/WPRS Bull., 18 (6) : 60-65.
Villemant C. & Ramzi H., 1995. Predators of gypsy moth (Lepidoptera : Lymantriidae) egg masses : spatio-temporal variation of their impact during the 1988-89 pest generation in the Mamora cork oak forest (Morocco). Entomophaga, 40 (3/4): 1-15.
Villemant C. & Andreï-Ruiz M.-C., 1999. Life-cycle and biological features of egg predators of Lymantria dispar (Lepidoptera : Lymantriidae) in the Mamora cork oak forest, Morocco. Eur. J. Entomol., 96 : 29-36.
Villemant C. & Fraval A., 1999. Les gradations de Lymantria dispar en Europe et en Afrique du Nord. IOBC/WPRS Bull. (sous presse)
Walnner W.E. & Walton G.S., 1979. Host defoliation: a possible determinant of gypsy moth population quality. Ann. entomol. Soc. Am., 72(1) : 62-67.
Vers
la page d'accueil d'OPIE-Insectes